
UASB中厌氧污泥脱铵及与硫酸盐还原的关系
运行期间,NH4+-N脱除量呈缓慢上升的趋势,而SO42--S的脱除量波动较大且呈不断下降趋势。在实验后期检测不到SO42--S的脱除,可能是由于污泥衰亡产生SO42-以及发生的硫自养反硝化反应,形成了硫的再循环。反应器前期出水S2-浓度较高、波动较大,之后S2-浓度逐渐稳定在1.45mg/L左右。由于在反应器中未检测到其他含硫物质,通过物料衡算得出至少存在35.06mg/L含硫物质积累于反应器中。
实验初期即观察到NH4+-N和SO42--S发生了脱除现象,但这一现象并不稳定。NH4+-N脱除量逐渐升高,SO42--S脱除量却逐渐降低。进水中存在的饱和溶解氧仅为8mg/L,按照硝化反应来计算不足以氧化等量的NH4+-N,但是NH4+-N仍有较高的脱除量,Zhang等人认为HCO3-有可能作为电子受体参与了N和S之间的反应,因此本研究考察了不同浓度的HCO3-对NH4+-N和SO42--S转化的影响。
2.2 HCO3-对NH4+-N和SO42--S转化的影响
本实验中添加NaHCO3以及KHCO3作为无机碳源维持反应器pH稳定,在连续流反应器持续运行至第162天时,NH4+-N、SO42--S及HCO3-浓度保持在100、60、305mg/L。如图3所示,分别在第187天、第209天和第237天提高HCO3-浓度至668、835、1002mg/L,发现NH4+-N脱除量逐渐上升,SO42-脱除量则未受到影响,在本实验中HCO3-并未参与到N和S的反应中。而反应器在第89天时通过批次实验也检测到3.4mgNH4+-N/gVSS的Anammox活性,即存在少量的Anammox菌。Anammox菌可通过厌氧氨氧化途径将氨氮转化后的少量NO2-作为电子受体,将氨氮氧化为N2,进一步提高氨氮的脱除量。
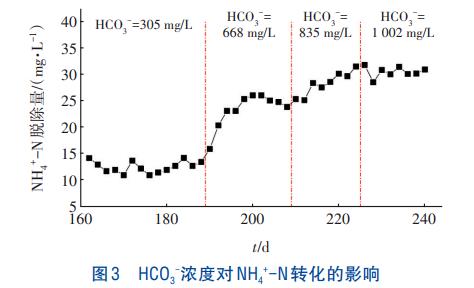
当HCO3-浓度提升至1002mg/L时,NH4+-N转化量趋于稳定。分析原因,一方面可能是反应器底部的Anammox菌较少,利用HCO3-的能力已达到饱和;另一方面可能是前期能够利用HCO3-的兼性自养微生物较少,导致HCO3-不能得到有效利用。对在反应器底部发现的黄色物质进行XRD检测,发现在转角为29.3°、35.9°、39.4°与43.1°处均有吸收峰,表明该物质主要为CaCO3。HCO3-大量积聚在反应器中,与进水中的Ca2+、Mg2+生成CaCO3及MgCO3等沉淀附着于颗粒污泥表面,不仅影响传质效率,还会加速颗粒污泥解体,进而影响出水水质。
此外,由于UASB反应器的特点,在长期运行过程中污泥层菌群在反应器内呈垂直分布,可能出现各污泥层生化反应相耦合,导致NH4+-N及SO42--S的脱除,而在本实验中NH4+-N与SO42--S的变化趋势也表明有其他电子受体及其他生化反应参与了NH4+-N和SO42--S的转化,因此检测不同高度出水水质,以探究可能的转化途径。
2.3 不同高度处NH4+-N与SO42-的转化情况
同一时刻在不同高度处采集水样进行检测,结果如图4所示。进水NH4+-N在反应器底部即被脱除,由100.34mg/L降至73.23mg/L。此外,部分厌氧颗粒污泥上浮裂解,细菌死亡,导致出水口处的NH4+-N浓度上升,这也与体系内检测到的ORP及TOC浓度变化一致。
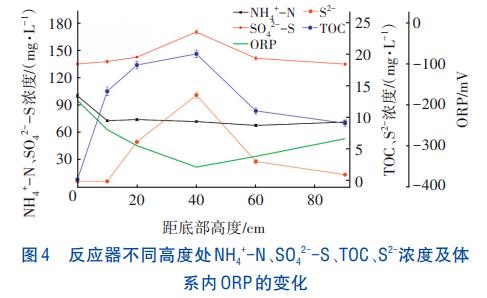
TOC及SO42--S浓度呈先上升后下降的趋势,SO42--S浓度先由135.16mg/L逐渐升至170.16mg/L,然后逐渐下降,在出水口处达到最低浓度135.08mg/L,由此观察到出水SO42--S未能脱除。在中部污泥层TOC浓度高达20.12mg/L,SRB菌利用大量有机物及SO42--S进行异养硫酸盐还原产生S2-,S2-浓度最高可达13.61mg/L。
由于进水中含有少量溶解氧,反应器底部发生了硝化作用使得NH4+-N浓度降低,而底部厌氧污泥死亡使TOC及SO42--S浓度逐渐升高。同时,厌氧污泥死亡时会释放H2O2及·OH等氧化性物质而氧化NH4+-N,导致NH4+-N在厌氧污泥层被脱除。硫自养反硝化反应与硝化反应相耦合,S0或S2-被重新氧化为SO42--S,在反应器中上层SRB菌利用SO42--S及TOC发生硫酸盐还原反应。综上所述,认为实验期间NH4+-N与SO42--S在不同位置分别被脱除。
2.4 反应器中N与S共同参与的反应
反应器底部NH4+-N由于硝化作用被氧化为NOx--N的同时,还存在硫酸盐还原的产物S2-及S0。在S0及NOx--N都存在的情况下,硫自养反硝化细菌逐渐被富集而发生反应,反应方程式如下:

批次实验2中分别添加S0以及NO2--N和NO3--N来证明反应器内发生的硫自养反硝化反应,对照组中仅放入与实验组等量未添加任何基质的污泥,空白组仅添加基质溶液,实验结果如图5所示。、

在厌氧条件下对照组中的微好氧菌死亡导致SO42--S浓度升高,实验组中NO2--N和NO3--N浓度下降,SO42--S浓度不断上升且远远高于对照组,说明反应器中可发生硫自养反硝化反应,而S0的再氧化很可能是后期SO42--S不发生脱除的原因之一。因此认为,污泥中的微生物可利用NOx--N与S0实现N元素与S元素在反应器中的协同循环。
2.5 反应器中的硫酸盐还原反应及其产物
运行初期SO42--S脱除量较高,之后逐渐降低,到后期SO42--S甚至不发生脱除。接种污泥浓度较高,MLVSS可达47.32g/L,在运行期间不断衰亡释放有机物。随着反应器的不断运行,污泥衰亡量逐渐降低,其释放的TOC浓度呈下降趋势,而SRB极有可能利用污泥衰亡释放的TOC进行异养硫酸盐还原,因此在厌氧环境下利用不同COD浓度进行批次实验以检测硫酸盐还原活性。
批次实验1中SO42--S浓度及ORP变化见图6。
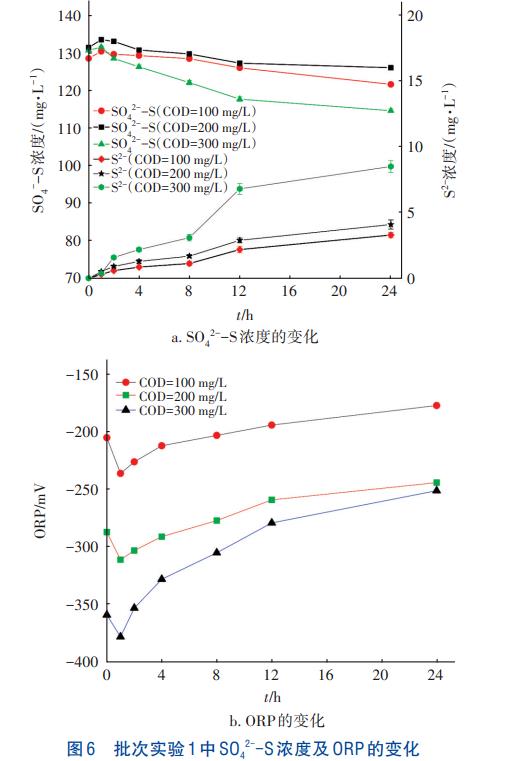
由图6可知,1h内SO42--S浓度均有不同程度的上升,原因可能是污泥中含有的微好氧菌死亡释放了部分SO42--S及有机物,而ORP则由于微生物死亡释放有机物而短暂下降。之后SRB菌利用有机物进行异养硫酸盐还原产生S2-,SO42--S浓度不断降低,ORP在COD被消耗后缓慢回升。在COD为300mg/L的条件下,测得硫酸盐还原活性最高,为9.91mgSO42--S/(gVSS·d)。
同时对反应器上层发现的部分黄色物质进行XRD检测,发现在转角为23.1°、25.9°、26.7°、27.7°及28.7°处均有吸收峰,显示有S0生成。由于S2-为耗氧型污染物,易被氧化物质氧化,而反应器上层由于漏氧、水封条件差或颗粒污泥上浮裂解释放氧化物质等因素,使得S2-逐渐被氧化为S0。
自来水以及污泥中所含Fe、Cu等金属元素容易与SRB还原生成的S2-结合生成含硫矿物,形成以金属或非金属物质作为晶核的沉淀,为此对底部黄色污泥进行SEM观察(如图7所示),同时对反应器底部及中部初期和后期污泥进行元素分析,结果见表2。从图7和表2可以看出,污泥中存在含硫沉淀形成的结晶物,且由于反应器内形成了CaCO3沉淀,反应器底部与中部污泥的Ca元素占比均明显提高。与初期相比,后期反应器底部污泥中的S元素占比较低,可能是由于发生硫自养反硝化导致S0重新生成了SO42--S。中部污泥的S元素则开始积累,S元素占比由初始的1.1%积累至2.3%,说明发生硫酸盐还原反应生成了S2-,之后被氧化为S单质积累在污泥层中,这也与2.3节的结果一致。

2.6 反应器中的氮和硫循环
反应器底部污泥负责NH4+-N的转化。一方面,实验初期接种的厌氧颗粒污泥中混有少量有机物,经过厌氧发酵后产生多种代谢产物,使异养菌大量增值同化NH4+-N,导致在厌氧环境下NH4+-N浓度降低;另一方面,由于进水桶未进行除氧,进水中携带少量的溶解氧(约8mg/L),当反应器底部兼性厌氧菌受到O2刺激时会产生H2O2,在反应器底部、中部及上部测得H2O2浓度分别为1.4、2.3、3.1mg/L,H2O2可氧化NH4+-N,提高NH4+-N的脱除量。而反应器内SO42--S的转化则主要利用体系内污泥死亡释放的TOC进行SO42--S还原。
当运行至第89天时,底部污泥逐渐由黑色转变为黄色,经批次实验证明底部污泥的硝化能力可达到30.25mgNH4+-N/(gVSS·d),且高通量测序同样表明,底部污泥中存在硝化菌。硝化菌利用少量溶解氧将NH4+-N转化为NO2--N,不仅为Anammox菌生长提供了所需的NO2--N,而且为Anammox菌增殖创造了条件。由于反硝化菌初始丰度比Anammox菌要高,对NO2--N的竞争力较强,初期Anammox菌的丰度较低,没有合适的底物可供利用,生长较为缓慢,导致NH4+-N脱除量较低。反应器经过长时间运行后,硝化活性逐渐增强,硝化菌以及H2O2等使得NH4+-N脱除量提高,提供更多的NO2--N从而促进Anammox菌的生长。第89天利用批次实验检测到反应器底部的Anammox活性为3.4mgNH4+-N/(gVSS·d),Anammox反应参与底部NH4+-N的转化,进一步提高了NH4+-N的脱除量。
在反应器运行至第150天时,SO42--S不发生脱除则是由于进水溶解氧使得厌氧污泥死亡,释放了额外的SO42--S。此外,由于污泥携带的有机物被消耗,SRB仅利用污泥衰亡产生的TOC进行SO42--S还原,SO42--S还原量逐渐减少。同时,在反应器底部发生的硫自养反硝化反应以及中部的硫氧化反应,也使得S2-或S0重新生成SO42--S,最终使得出水SO42--S基本不发生转化。
因此推断,在反应器下层发生的是Anammox、硝化及硫自养反硝化耦合的生化反应,在中层与上层则发生的是硫酸盐还原及硫氧化反应,见图8。
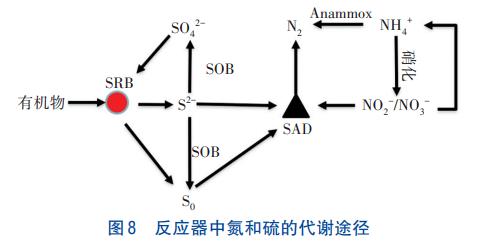
2.7 微生物菌群解析
利用高通量技术分析了反应器在0、89、240d时污泥的微生物菌群结构,结果见图9。可知,在反应器的3个运行阶段,硝化菌、反硝化菌、硫酸盐还原菌、硫自养反硝化菌以及硫氧化菌均存在。
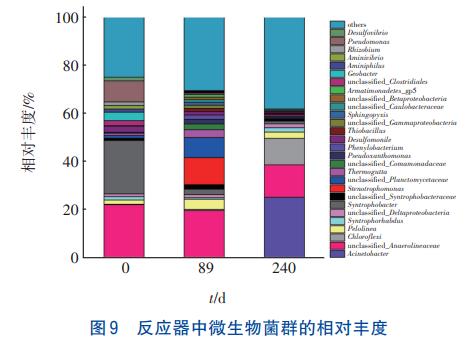
在反应器底部检测到Acinetobacter,其丰度由初始的0.023%增至第240天的25.22%,同时检测到Comamonadaceae、Pseudoxanthomonas、Thiobacillus、Thermomonas等反硝化菌,初始总丰度仅为0.5%,在第89天提高至6.51%。其中,Thiobacillus是一种硫自养反硝化菌,其丰度由初始的0逐渐提高至1.32%,说明反应器中确实存在硫自养反硝化反应,这与2.5节的结果相吻合。高通量测序中并未发现Anammox菌,但在反应器运行至第89天时利用批次实验检测到较低的Anammox活性。反应器中Desulfomonile、Desulfovibrio及Syntrophobacteraceae等SRB菌,在初始接种污泥中的总丰度可达到4.96%,随着反应器内有机物的减少,SRB菌的总丰度呈下降趋势,在第240天下降至1.67%,这也与反应器中硫酸盐脱除量降低的趋势相吻合。同时,在反应器中存在少量的硫氧化菌属,例如Rhodopseudomonas、Rhodobacteraceae等,可利用H2S进行代谢自养生长,总丰度可达到0.188%。
在第162天提高HCO3-浓度以后,NH4+-N脱除量增加。在主要负责脱氮的底部污泥中,检测到Acinetobacter菌的丰度提高至78.87%。有研究者发现,Acinetobacter菌具有硝化及反硝化能力,甚至部分Acinetobacter菌群可进行异养硝化脱氮。底部污泥衰亡产生的少量有机物为Acinetobacter菌群异养硝化提供了基质,同时进水中少量的DO以及H2O2为Acinetobacter的好氧反硝化提供了条件,导致其大量增殖,反应器中氨氮的脱除主要发生在底部污泥中,而其与混合污泥菌群的差别仅在于Acinetobacter的丰度。即Acinetobacter的丰度决定了反应器的脱氮效果,因此认为Acinetobacter在反应器中起主要的脱氮作用。Desulfomonile、Desulfovibrio及Syntrophobacteraceae则是反应器中主要的SRB菌,利用SO42-作为电子受体进行SO42-还原反应。Thiobacillus作为反应中的硫自养反硝化细菌,可将SO42-还原生成的S2-或S0以NOx--N为电子受体氧化为SO42-,与硫氧化菌共同形成硫的再循环,其丰度逐渐提高使得SO42-的脱除量进一步降低,这也与2.1节中的推测相吻合。在本实验中并未发现Cai等人鉴定出的可以同时利用NH4+-N以及SO42--S的BacillusBenzoevorans菌株,也未曾发现Liu等人在Anammox反应器中分离得到的脱氮除硫功能菌种Anammoxoglobussulfate。
因此,在本研究的反应器中,硝化、Anammox、异养反硝化及硫自养反硝化共同构成了系统内的氮循环;硫酸盐还原、硫自养反硝化及硫好氧氧化共同构成了系统内的硫循环,氮、硫循环耦合使得反应器中出现硫酸盐还原厌氧氨氧化。
3、结论
①改变UASB反应器进水N/S值时,NH4+-N与SO42--S的脱除量并未受到明显影响。体系内存在的Anammox反应、兼性厌氧微生物产生的H2O2以及提高HCO3-浓度均有助于NH4+-N脱除量的提高。在将HCO3-浓度提高至1002mg/L后,NH4+-N脱除量不再升高。
②由于UASB反应器的特点,在不同高度污泥层形成了不同种类的微生物菌群,NH4+-N和SO42--S在不同位置分别被脱除,由不同层的氮、硫生化反应相互耦合形成硫酸盐型厌氧氨氧化现象。
③溶解氧使得厌氧污泥死亡产生额外的SO42--S,SO42--S还原生成的S2-被Rhodopseudomonas、Rhodobacteraceae等硫氧化菌以及硫自养反硝化菌重新氧化为SO42--S,在反应器内部形成硫循环,导致出水SO42--S不发生脱除。而Acinetobacter主要负责反应器内NH4+-N的脱除,与Anammox、异养反硝化及硫自养反硝化共同构成体系内的氮循环。
建树环保是一家专注于水污染治理、大气污染治理及环境专用设备制造与销售的建设工程施工企业,建树环保持有环保工程专业承包二级资质和安全生产许可证,为客户从“方案设计”、“工程施工”、“安装调试”、“售后服务”环节提供服务。联系电话:135 5665 1700。 






